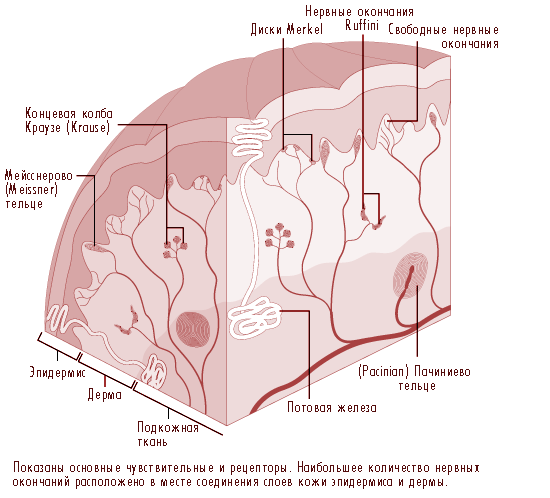
Рис. 11.19 Схематическая иллюстрация поперечного среза кожного покрова
Система кожного восприятия обладает теми же основными элементами, что и все основные системы органов чувств. Свойства внешнего мира, такие как цвет, звук или вибрация принимаются специальными окончаниями нервных клеток, которые называются чувствительными рецепторами и преобразуют данные из внешнего мира в нервные импульсы. Эти сигналы затем передаются в центральную нервную систему, где становятся основой для восприятия окружающего нас мира.
Необходимо понимать три основных принципа этих процессов. Первый заключается в том, что энергия и изменения в уровнях энергии могут быть восприняты лишь чувствительным органом, предназначенным для обнаружения специфического типа энергии. Именно поэтому микроволны, рентгеновские лучи и ультрафиолет являются опасными; мы не приспособлены к тому, чтобы обнаруживать эти излучения, так что даже на уровнях, опасных для жизни, они не могут быть восприняты. Второй принцип заключается в том, что наши восприятие, несомненно, являются несовершенным отражением действительности, поскольку наша центральная нервная система ограничена отражением только тех сигналов, которые переданы ей чувствительными рецепторами. Третий принцип состоит в том, что наши системы органов чувств обеспечивают нам информацию об изменениях в окружающей среде более точно, чем о ее статических состояниях. Мы снабжены рецепторами, чувствительными к мерцанию света, например, или к незначительным изменениям температуры, вызванным легким бризом; но гораздо меньше приспособлены к тому, чтобы получать информацию, скажем, о постоянной температуре или постоянном давлении на кожный покров.
Традиционно кожные ощущения разделяют на две категории: поверхностные и глубокие. Глубокая чувствительность основана на рецепторах, расположенных в мышцах, сухожилиях, связках и надкостнице (оболочка окружающая кости); поверхностная кожная чувствительность, о которой здесь пойдет речь, обеспечивает нас информацией, полученной многочисленными, различными по строению нервными окончаниями, расположенными в эпидермисе и дерме.
Все чувствительные нервы, связывающие кожные рецепторы с центральной нервной системой, имеют приблизительно одинаковое строение. Группа больших тел нервных клеток, образуют нервный узел - ганглий, расположенный рядом со спинным мозгом и соединенный с ним посредством узкого ответвления клеточного ствола, называемого нейритом. Большинство нервных клеток или нейронов, которые начинаются у спинного мозга, распространяют свои нейриты на кости, мышцы, связки или, как в случае с кожной чувствительности, на кожный покров. Точно так же как обычный изолированный провод, каждый нейрит на всем своем протяжении и на окончаниях покрыт защитными слоями клеток известных как клетки Schwann. Эти клетки Schwann вырабатывают вещество, известное как миелин, которое окутывает нейрит подобно оболочке. Крошечные отрезки нейрита по всей его длине незаполненные миелином известны в качестве узлов Ренвьера. И, наконец, на конце нейрита находятся образования, специализирующиеся на получении и повторной передаче информации из внешней среды: чувствительные рецепторы (Mountcastle 1974).
Кожные рецепторы, также как и все остальные чувствительные рецепторы, классифицируются по двум признакам: анатомическое строение рецептора и тип электрических сигналов, которые они посылают по нервным волокнам. Рецепторы определенной структуры обычно называются по имени ученых их открывших. Относительно небольшое количество классов чувствительных рецепторов, обнаруженных в кожном покрове, может быть разделено на три основные категории: механорецепторы, терморецепторы и ноцицепторы.
Все эти рецепторы могут передавать информацию о специфических стимулах только после того, как она зашифрована в определенный тип электрохимического нейроязыка. Это нервные коды используют изменяющиеся частоты и модели нервных импульсов, которые ученые только что начали расшифровывать. В действительности значительная часть нейрофизиологических исследований полностью посвящена изучению чувствительных рецепторов и механизмов, при помощи которых они переводят энергетические состояния окружающей среды в нервные коды. Как только код создан, он передается в центральную нервную систему по центростремительным волокнам - нервным клеткам, которые обслуживают рецепторы, передавая сигналы в центральную нервную систему.
Сообщения, составляемые рецепторами, могут быть классифицированы на основе реакции, получаемой на непрерывную, стабильную стимуляцию: медленно адаптирующиеся рецепторы посылают электрохимические импульсы в центральную нервную систему в течение действия постоянного стимула, в то время как быстро адаптирующиеся рецепторы постепенно снижают свою активность в этом направлении в присутствии устойчивого стимула, пока не достигают нижнего базового уровня либо не замолчат полностью, таким образом, прекращая информировать центральную нервную систему о продолжающемся присутствии стимула.
Таким образом, различные ощущения, такие как боль, тепло, холод, давление и вибрация, создаются деятельностью определенных классов чувствительных рецепторов и связанных с ними нервных волокон. Термины “трепет” и “вибрация”, например, используются для обозначения двух мало отличающихся друг от друга вибрационных ощущений, которые шифруются двумя различными классами чувствительных к вибрации рецепторов (Mountcastle et al. 1967). Каждая из трех основных категорий ощущения боли, известных как покалывание, жгучая боль и ломящая боль, связана с определенным классом центростремительных волокон ноцицептора. Однако нельзя говорить о том, что отдельное ощущение обязательно затрагивает лишь один вид рецепторов; каждое отдельное ощущение может вызываться несколькими видами рецепторов, и, в реальности, ощущения могут отличаться друг от друга соотношением долевого участия различных видов рецепторов в их производстве (Sinclair 1981).
Вышеизложенное основано на особой гипотезе, впервые сформулированной для чувствительной функции кожи немецким врачом по имени Фон Фрей в 1906г. Хотя в течение последнего столетия были предложены, по крайней мере, две других теории, которые являются такими же или более популярными, чем теория Фон Фрея, однако его гипотеза на данный момент основательно подтверждается фактическими данными.
Рецепторы, реагирующие на постоянное давление на кожный покров
В кистях рук расположены относительно большие миелиновые волокна (от 5 до 15 mm в диаметре), которые берут свое начало в подкожной нервной сети, так называемом субпапиллярном нервном сплетении, и заканчиваются разветвлением нервных окончаний в области соединения дермы и эпидермиса (рисунок 11.19). В покрытом волосами кожном покрове эти нервные окончания образуют видимые поверхностные структуры известные как сосочки, (купола соприкосновения); в кожном покрове, лишенном волос, нервные окончания находятся у основания гребешков кожи - тех, что формируют отпечатки пальцев. В куполе соприкосновения каждый кончик нервного волокна, или нейрит, окружен специальной эпителиальной клеткой известной как клетка Меркеля (см. рисунки 11.20 и 11.21).
--------------------------------------------------------------------------------
Рис. 11.19 Схематическая иллюстрация поперечного среза кожного покрова
--------------------------------------------------------------------------------
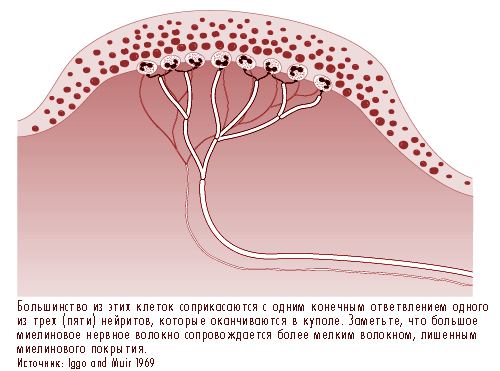
Рис. 11.20 Купол прикосновения в каждой из гребешков кожного покрова содержит от 30 до 70 клеток Merkel
--------------------------------------------------------------------------------
Рис. 11.21 Большее увеличение, полученное при помощи электронного микроскопа. Видно соединение клетки Merkel,
специальной эпителиальной клетки, с базальной мембраной, которая отделяет эпидермис от дермы
--------------------------------------------------------------------------------
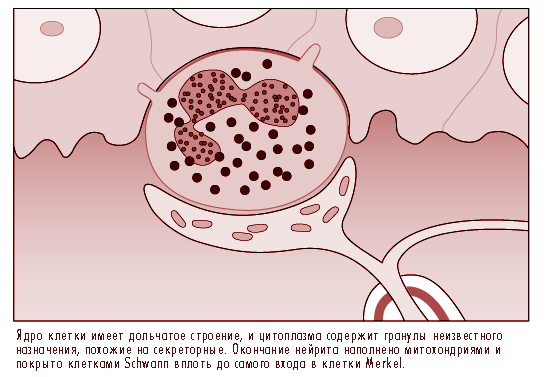
Нейронный комплекс клетки Меркеля преобразует механическую энергию в нервные импульсы. Хотя на данный момент о роли клетки или ее механизме преобразования известно немного, она была идентифицирована в качестве медленно адаптирующегося рецептора. Это означает что давление, производимое на сосочки (купол соприкосновения), содержащий клетки Меркеля заставляет рецепторы производить нервные импульсы в течение всего периода действия стимула. Частота этих импульсов возрастает в определенной пропорции к интенсивности стимула, таким образом, информируя мозг о продолжительности и силе давления на кожный покров.
В кожном покрове имеется еще один медленно адаптирующийся рецептор (подобно клетке Меркеля), который передает импульсы в ЦНС о силе и продолжительности постоянного давления на кожный покров. Видимый только через микроскоп, этот рецептор, известный как рецептор Руффини, состоит из группы нейритов берущих свое начало из миелинового волокна и окруженных клетками соединительной ткани. Внутри капсулы расположены волокна, которые, по-видимому, передают сигналы о локальных кожных деформациях нейритам, которые в свою очередь передают сообщения в центральную нервную систему. Давление на кожный покров вызывает процесс непрерывного производства нервных импульсов; как и в случае с клетками Меркеля, частота нервных импульсов пропорциональна интенсивности стимула.
Несмотря на свою схожесть, клетки Меркеля имеют значительное отличие от рецепторов Руффини. В то время как при стимуляции рецепторов Руффини возникает ощущение, стимуляция куполов соприкосновения, вмещающих клетки Меркеля, не вызывает сознательного ощущения; купол соприкосновения, таким образом, является неизвестным рецептором, поскольку его фактическая роль в нервной функции не выяснена. Рецепторы Руффини в настоящее время считаются единственными рецепторами, способными обеспечить нервную систему импульсами, необходимыми для восприятия давления или постоянного соприкосновения с кожей. Кроме того, как было продемонстрировано, медленно адаптирующиеся рецепторы Руффини дают человеку возможность определять величину давления на кожный покров в соответствии с его интенсивностью.
Рецепторы, реагирующие на вибрацию и движение кожного покрова
В отличие от медленно адаптирующихся механорецепторов, быстро адаптирующиеся рецепторы молчат в процессе устойчивого состояния кожного покрова. Однако они хорошо приспособлены к работе с сигнальной вибрацией и перемещением кожного покрова. Были выделены две основные категории подобных рецепторов: рецепторы, расположенные в покрытом волосами кожном покрове и связанные с отдельными волосками; и рецепторы, которые формируют капсулированные (чувствительные) нервные окончания в кожном покрове, лишенном волос.
Рецепторы, обслуживающие волосы
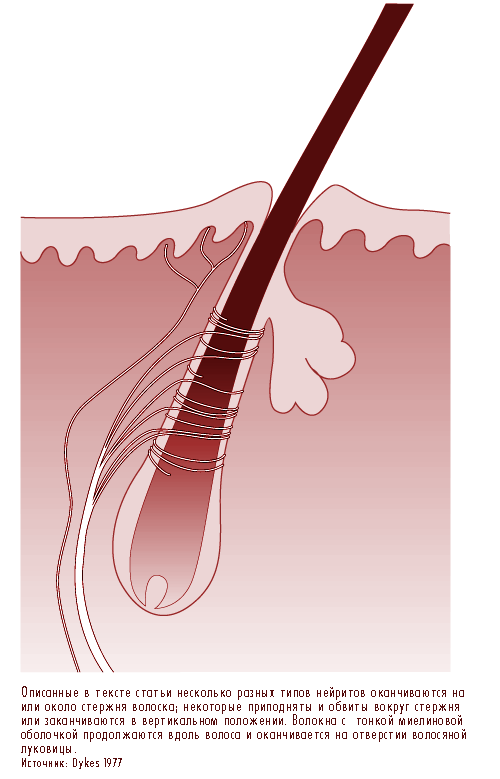
Типичный волосок окружен сетью нервных окончаний, из которой выходит от пяти до девяти больших миелиновых нейритов (рисунок 11.22). У приматов эти окончания разделяются на три категории: ланцетовидные окончания, шпинделеподобные окончания и сосковидные окончания. Все три являются быстро адаптирующимися рецепторами, так что постоянное отклонение волосков вызывает нервные импульсы только непосредственно во время перемещения. Таким образом, эти рецепторы особо чувствительны к перемещению или вибрации, однако дают недостаточно информации о давлении или постоянном соприкосновении либо вообще не поставляют таковой.
--------------------------------------------------------------------------------
Рис. 11.22 Стержни волосков являются платформой для нервных окончаний, отвечающих за обнаружение движений
--------------------------------------------------------------------------------
Копьевидные окончания появляются из сильно миелинизированного волокна, которое формирует сеть вокруг волосков. Нейриты нервных окончаний лишаются своего обычного покрытия клеток Schwann и прокладывают себе путь между клетками в основании волоска.
Шпинделеподобные окончания сформированы из окончаний нейритов окруженных клетками Schwann. Нервные окончания подходят к покатому стержню волоска и оканчиваются в полукруглом пучке сразу под сальной железой. Сосковидные окончания отличаются от шпинделеподобных тем, что вместо того, чтобы оканчиваться на стержне волоска, они завершаются в качестве свободных нервных окончаний окружающих волосковое отверстие.
Предположительно, среди разных типов рецепторов, находящихся на волосках, имеются функциональные различия. Частично это может быть отнесено к структурным различиям (к тому, каким образом нервы оканчиваются на стержне волоска), частично - к разнице в диаметре нейритов, так как нейриты разных диаметров связаны с разными релейными областями. На данный момент функции рецепторов покрытого волосами кожного покрова остаются предметом изучения.
Рецепторы кожного покрова, не покрытого волосами
Взаимозависимость между анатомическим строением рецептора и нервными импульсами, которые он вызывает, наиболее очевидна у крупных и легко управляемых капсулированных (чувствительных) нервных окончаниях в кожном покрове, лишенном волос.
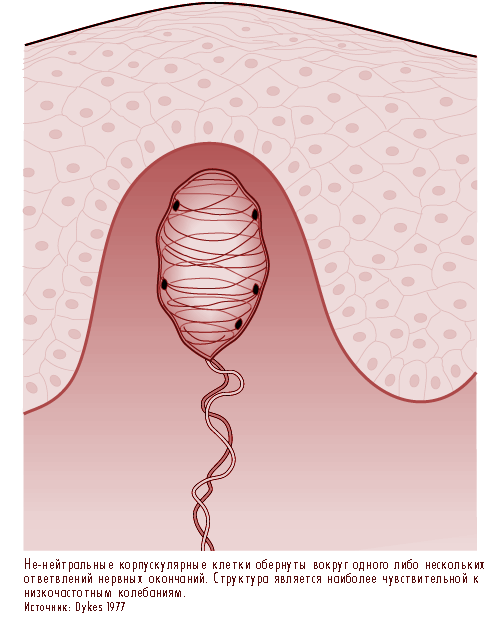
Наиболее хорошо понятыми на данный момент являются пачиниевы (pacininan) и мейсснеровы тельца (Meissner), которые, подобно вышеописанными нервным окончаниям кожи с волосяным покровом, сообщают ощущения вибрации.
Пачиниевы тельца (Pacinian) достаточно велики для того, чтобы быть видимыми невооруженным глазом, что облегчает ассоциацию рецептора с конкретной нервной реакцией. Эта луковицеподобная структура, расположенная в дерме, обычно вокруг сухожилий или связок, имеет размер в 0.5 х 1.0 мм. С ней связано одно из самых больших центростремительных волокон организма, имеющее диаметр от 8 до 13 ![]() m и проводимость от 50 до 80 метров в секунду. Ее анатомия хорошо изучена при помощи оптических и электронных микроскопов.
m и проводимость от 50 до 80 метров в секунду. Ее анатомия хорошо изучена при помощи оптических и электронных микроскопов.
Основным компонентом тельца является внешний сердечник, сформированный из клеток, заполненных жидкостью. Внешний сердечник сам окружен оболочкой, которая пронизана центральным каналом и сетью капилляров. Через канал проходит единственное миелиновое нервное волокно диаметром от 7 до 11 ![]() м, которое затем становится длинным, немиелиновым нервным окончанием, проникающим глубоко в центр тельца. Нейрит нервного окончания имеет эллиптическую форму и снабжен ветвеобразными отростками.
м, которое затем становится длинным, немиелиновым нервным окончанием, проникающим глубоко в центр тельца. Нейрит нервного окончания имеет эллиптическую форму и снабжен ветвеобразными отростками.
Пачиниевы тельца является быстро адаптирующимся рецептором. Таким образом, подвергаясь воздействию постоянного давления, она производит импульсы только при начале и окончании стимуляции. Она реагирует на высокочастотные колебания (от 80 до 400 Гц) и наиболее чувствительна к колебаниям с частотой примерно 250 Гц. Часто эти рецепторы реагируют на колебания, передаваемые по костям и сухожилиям, и вследствие своей чрезвычайной чувствительности, они могут быть активированы, стоит вам всего лишь подуть на ладонь (Martin 1985).
В дополнение к Пачиниевым тельцам в безволосом кожном покрове имеется другой быстро адаптирующийся рецептор. Большинство исследователей полагает, что это мейсснеровы тельца, расположенные в сосочках кожного покрова. Чувствительный к низкочастотным колебаниям от 2 до 40 Гц, этот рецептор состоит из ответвлений нервных окончаний среднего размера миелинового нервного волокна, окруженного одним либо несколькими слоями клеток, которые похожи на измененные шванновы клетки (Schwann) и называются ламинарными клетками. Нейриты и ламинарные клетки рецептора могут быть связаны с базальной клеткой в эпидермисе (рисунок 11.23).
--------------------------------------------------------------------------------
Рис. 11.23 Тельца Meissner представляет собой закрытый в просторную капсулу чувствительный рецептор в кожном
бугорке безволосой кожи
--------------------------------------------------------------------------------
Мейсснеровы тельца выборочно инактивируются при помощи инъекции местного обезболивающего через кожу, ощущение трепета или низкочастотной вибрации при этом теряется. Это говорит о том, что Мейсснеровы тельца функционально дополняет высокую частотную способность Пачиниевых телец. Вместе эти два рецептора обеспечивают возникновение и передачу нервных импульсов, достаточных для того, чтобы обеспечивать человеку чувствительность к вибрации всех диапазонов (Mountcastle et al. 1967).
Кожные рецепторы, связанные со свободными нервными окончаниями
В дерме находится также большое количество как миелиновных, так и лишенных миелинового покрытия нервных волокон, назначение которых еще не установлено. Многие из них только проходят через дерму на своем пути к поверхностному слою кожи, мышцам или надкостнице, в то время как другие (как миелиновые, так и лишенные миелинового покрытия), похоже, оканчиваются в дерме. За некоторым исключением, типа пачиниевых телец, до сих пор в недостаточной степени определено, каким образом оканчивается большинство нервных волокон, такие волокна просто причисляют к свободным нервным окончаниям.
В то время как для дифференциации этих недостаточно изученных нервных окончаний необходимо проведение дополнительных анатомических исследований, при помощи физиологического исследования специалисты пришли к заключению, что эти волокна кодируют разнообразные сигналы окружающей среды. Например, свободные нервные окончания, которые расположены на границе между эпидермисом и дермой отвечают за кодирование таких сигналов окружающей среды, как холод, тепло, боль, зуд, и щекотка. На данный момент не известно, какие из разнообразных классов этих небольших волокон передают каждое из этих ощущений индивидуально.
Кажущаяся анатомическая схожесть этих свободных нервных окончаний вызвана скоре всего ограниченностью наших исследовательских методов, так как мы медленно начинаем замечать структурные различия между свободными нервными окончаниями. Например, в кожном покрове, лишенном волос, были определены две различных модели окончания свободных нервных окончаний: толстая, короткая модель и длинная и тонкая. Исследования человеческого кожного покрова, покрытого волосками, выявили распознаваемые гистохимическим путем нервные окончания, которые оканчиваются на границе между дермой и эпидермисом: penicillate и сосочковые окончания. Первые выходят из лишенных миелинового покрытия волокон и формируют сеть окончаний; последние напротив, являются продолжением миелиновых волокон и, как было упомянуто выше, заканчиваются вокруг отверстий волосков. Возможно, эти структурные различия объясняют различия в функциях.
Хотя на данный момент остается невозможной дифференцировка специфических функций отдельных структурных единиц, из физиологических экспериментов становится ясным, что существуют категории свободных нервных окончаний, имеющих функциональные различия. В частности, было обнаружено миелиновое нервное волокно, ответственное за восприятие человеком ощущения холода. Другое нервное волокно, лишенное миелинового покрытия, обслуживающее свободные нервные окончания, реагирует на тепло. Каким образом один класс свободных нервных окончаний выборочно реагирует на снижение температуры, в то время как другой класс реагирует на увеличение температуры кожного покрова, до сих пор остается неизвестным. Исследования показывают, что возбуждение одного небольшого волокна со свободным окончанием может быть ответственно за восприятие человеком зуда или щекотки, тогда как два класса небольших нервных волокон особенно чувствительны к повреждающим механическим, химическим или тепловым воздействиям, формируя в ЦНС ощущения покалывания или жгучего болевого ощущения (Keele 1964).
Неполные знания о зависимости между анатомическим строением и физиологической реакцией создает трудности в управлении и контроле за такими нарушениями, как каузальгия, парестезия и гиперпатия, которые на сегодняшний день ставят врачей в затруднительное положение. Поэтому необходимо дальнейшее изучение данной проблемы.
Поражение периферического нерва
Согласно представлениям ученых, существуют нервные функции двух категорий: сенсорная и моторная. Поражение периферического нерва, обычно вызываемое сдавливанием либо разрывом нерва, может привести к расстройству одной либо обеих нервных функций, причем это зависит от типа волокон пораженного нерва. Некоторые аспекты потери моторной функции, похоже, были неверно истолкованы либо упущены из виду, поскольку эти сигналы не поступают к мышцам, но скорее воздействуют на автономный контроль кровеносных сосудов, температурный режим, природу и толщину эпидермиса, и состояние кожных механорецепторов. Данная потеря моторной иннервации не является предметом обсуждения в этой статье, также мы не будем затрагивать тему потери иннервации, приводящей к поражению органов чувств помимо тех, что отвечают за кожные ощущения.
Потеря сенсорной иннервации усиливает уязвимость кожи к последующим поражениям, так как способствует возникновению областей, нечувствительных к боли и соответственно неспособных передать в ЦНС информацию о потенциально опасных воздействиях. После нанесения повреждения процесс восстановления потерявшего болевую чувствительность кожного покрова протекает чрезвычайно медленно, возможно частично из-за отсутствия автономной иннервации, которая обычно управляет такими ключевыми факторами как температурный режим и питание клетки.
Лишенные иннервации в течение нескольких недель кожные чувствительные рецепторы начинают атрофироваться, процесс, который легко наблюдать на примере покрытых капсулой рецепторов типа пачиниевых (pacininan) и мейсснеровых телец (Meissner), Вслед за восстановлением нейрита, если это возможно, может восстановиться и функция, однако ее качественные характеристики будут находиться в прямой зависимости от природы исходной травмы и от продолжительности периода нарушения иннервации (McKinnon and Dellon 1988).
Восстановление, следующее за поражением нерва при дробном поражении, происходит быстрее и является намного более полным и более функциональным, чем восстановление после разрыва нерва. Два фактора объясняют более благоприятные прогнозы на выздоровление после дробного поражения нерва. Во-первых, в данном случае большее количество нейритов может снова вступить в контакт с наружным слоем, чем после поперечного рассечения нерва; во-вторых, связи с исходной областью восстанавливаются под управлением шванновых клеток (Schwann) и прокладками, известными в качестве базальной мембраны, причем обе эти структуры остаются невредимыми при дробном поражении нерва, тогда как после поперечного рассечения нервы часто перемещаются на другие области кожного покрова, следуя по путям, проложенным другими шванновыми клетками. В последнем случае в соматосенсорную кору головного мозга направляется искаженная пространственная информация. В обоих случаях, однако, восстанавливающиеся нейриты способны найти путь к сенсорным рецепторам того же класса, к которому они относились до этого.
Процесс восстановления иннервации кожного рецептора проходит постепенно. Когда растущий нейрит достигает поверхности кожного покрова, поля восприятия по размерам уступают обычным, в то время как порог восприятия превосходит обычный. Эти рецептивные точки со временем расширяются и постепенно объединяются в большие поля. Чувствительность к механическим стимулам становится больше и часто приближается к чувствительности нормальных чувствительных рецепторов данного класса. Исследования воздействия постоянного соприкосновения, перемещающегося соприкосновения и вибрации показало, что сенсорная модальность, присущая различным типам рецепторов, восстанавливается в областях, потерявших болевую чувствительность (области кожного покрова с поврежденным периферическим нервом), с различной скоростью.
При рассмотрении под микроскопом лишенная иннервации кожа без волосяного покрова кажется тоньше обычной, имеет более плоские папиллярные линии и меньшее количество клеточных слоев. Это подтверждает, что нервы выполняют также трофическую функцию по отношению к коже. Вскоре после восстановления иннервации, папиллярные линии становятся более развитыми, эпидермис утолщается, и можно обнаружить нейриты пронизывающие базальную мембрану. По возвращении нейрита в мейсснерово тельце, оно начинает расти, и атрофированные до этого структуры начинают принимать исходную форму. Если нарушение иннервации носило продолжительный характер, рядом с атрофированным тельцем может сформироваться новое (Dellon 1981).
Как можно заметить, для понимания последствий поражения периферического нерва необходимы знания о нормальном процессе его функционирования, также как и о степени функционального восстановления. В то время как для одних нервных клеток кожного покрова такая информация является доступной, другие клетки до сих пор не изучены, оставляя нас в неведении об их роли в жизнедеятельности организма.